Organites, ubiquitinylation et signalisation
Thèmatique de recherche
Nos projets de recherche actuels se focalisent désormais principalement sur le rôle de l'ubiquitination non dégradative (ubiquitination dite en K63), mais également sur les dé-ubiquitinations, dans les voies de signalisation conduisant notamment à l'activation des facteurs de transcription NF-?B ou bien IRF3. De plus, nous nous intéressons au rôle d'organites intracellulaires comme le réticulum endoplasmique, les mitochondries ou l'Appareil de Golgi dans les voies de signalisation conduisant à l'activation de ces facteurs de transcription.

projet 1
Implication du réticulum endoplasmique dans l'intégration du signal externe conduisant à l'activation de NF-κB
L'immunité innée et adaptative engagent la signalisation conduisant à l'activation de NF-κB par l'ubiquitination sélective d'adaptateurs spécifiques du contexte. Nous avons observé que ces Transmetteurs Ubiquitinylés Conduisant à l'Activation de NF-κB (que l'on appellera ici les TUCAN), c'est à dire RIP1, Bcl-10, TRAF2, RIP2 ou bien IRAK1 s'accumulent dans la fraction contenant les membranes lourdes (mitochondries, réticulum endoplasmique, golgi, lysosomes etc) et plus particulièrement au niveau du côté cytosolique de la membrane du réticulum endoplasmique (RE) suite à la stimulation respectivement du TNFR, TCR, CD40, Nod2, TLR4, TLR3 et RLR (Alexia et al., 2013).
Il est intéressant de noter que dans un système in vitro, nous avons observé que la fraction de membranes lourdes (qui, rappelons-le, contient notamment le RE) de cellules activées est capable d'induire la phosphorylation de IκBα, un des inhibiteurs de NF-κB. Ceci confirme que le RE, via l'accumulation des TUCAN, contient alors l'information nécessaire pour permettre l'activation de NF-κB.
Par spectrométrie de masse, nous avons identifié la méthadérine (MTDH), une protéine ancrée au RE, comme un partenaire des TUCAN. L'extinction de MTDH diminue sélectivement l'activation de NF-κB suite à différents stimuli, notamment le TNFα et de fait inhibe la production de cytokine ou bien l'activation des lymphocytes (Alexia et al., 2013).
Bien que les TUCAN sont normalement ubiquitinylés suite aux diverses stimulations, nous avons observé que moins de TUCAN s'accumulaient au RE dans les cellules déficientes en MTDH, suggérant que la MTDH permet l'accumulation des TUCAN au RE, ce qui propage ultérieurement la signalisation NF-κB (Alexia et al., 2013).
Nos résultats impliquent donc que la MTDH intègre les signaux en amont pour les convertir en réponses biologiques à travers l'axe IKK/NF-κB (Figure 1) (Alexia et al., 2013).
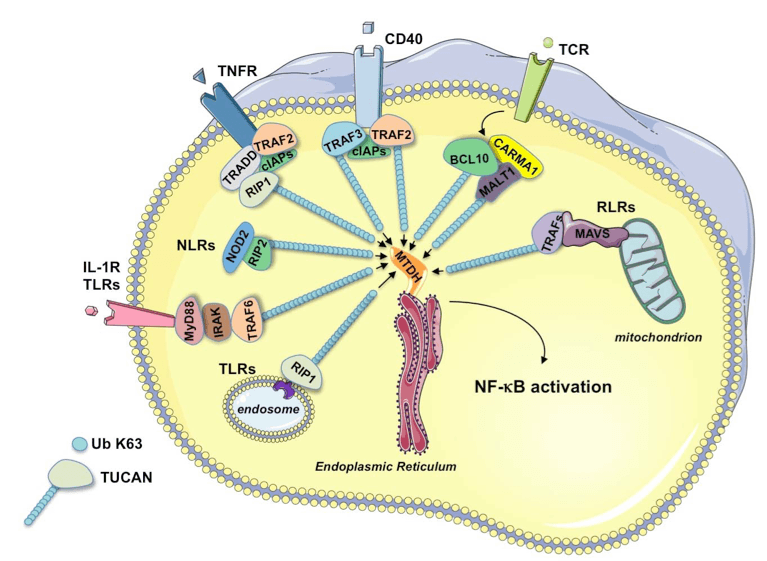
Figure 1. La MTDH fonctionne comme une clef de voute pour reconnaître les TUCAN et les cristalliser à la surface du RE
A l'aide de protéines recombinantes, nous avons montré que la MTDH se lie essentiellement aux chaines d'ubiquitine K63 (Alexia et al., 2013). Nous pensons ainsi que la MTDH pourrait agir comme un « senseur » des protéines ubiquitinylées. La petite quantité d'IKK et d'IκBα observée dans les membranes lourdes suggère que le « methadherinosome » constitue une étape importante dans le processus de l'activation de NF-κB.
D'autres études sont nécessaires pour comprendre précisément comment les signalosomes émanant des immuno-récepteurs maturent puis transitent via le RE pour finalement s'associer et activer NF-κB via le complex IKK. Nous disposons désormais de cellules issues de souris déficientes en MTDH pour répondre à ces questions..
Une question passionnante est quel peut être le rôle du RE dans la transduction du signal NF-κB via la MTDH ancrée à cet organite. Il est en effet fort probable que si la MTDH n'est pas associée au RE, alors la signalisation sera affectée. Nous sommes en train de vérifier cela. De la même manière, MAVS, protéine essentielle à la transduction du signal après activation des RLRs, est ancrée à la membrane mitochondriale externe et doit être absolument localisée à cet endroit pour pouvoir exercer sa fonction, sans que l'on ne sache pour autant encore quel est vraiment le rôle de la mitochondrie. De plus, nous essayons de comprendre si l'expression de la MTDH est régulée, notamment par les nombreuses E3 ubiquitine ligases associée au RE.

Organigramme de l’équipe
| Nom | Position | Institution | Thématique principale |
| ARNOULT damien | CR1 | CNRS | |
| SILVA DA COSTA Leandro | Post-doc | ||
| ZEMIRLI Naima | Post-doc | ||
| POURCELOT Marie | Doctorante | Paris XI | |
| RODERS Nathalie | Doctorante | Paris XI | |
| OUTLIOUA Ahmed | Doctorante | Univ. Casablanca | |
| GILBERT Sophie | Etudiante M2 | Paris XI | |
| LOYANT Roxane | Etudiante BTS | ||
| PORTIER | Technicien | Paris XI |
Publications marquantes, Brevets...
2014
-Zemirli N, Pourcelot M, Dogan N, Vazquez A, Arnoult D. The E3 ubiquitin ligase RNF121 is a positive regulator of NF-?B activation. Cell Commun Signal. 2014 Nov 12;12(1):72. [Epub ahead of print]
-Pourcelot M, Arnoult D. Mitochondrial dynamics and the innate antiviral immune response. FEBS J. 2014 Sep;281(17):3791-802.
-Soares F, Tattoli I, Rahman MA, Robertson SJ, Belcheva A, Liu D, Streutker C, Winer S, Winer DA, Martin A, Philpott DJ, Arnoult D, Girardin SE. The mitochondrial protein NLRX1 controls the balance between extrinsic and intrinsic apoptosis. J Biol Chem. 2014 Jul 11;289(28):19317-30
-Zemirli N, Pourcelot M, Ambroise G, Hatchi E, Vazquez A, Arnoult D. Mitochondrial hyperfusion promotes NF-κB activation via the mitochondrial E3 ligase MULAN. FEBS J. 2014 Jul;281(14):3095-112
2013
-Alexia C, Poalas K, Carvalho G, Zemirli N, Dwyer J, Dubois S, Hatchi E, Cordeiro N, Smith SS, Castanier C, Le Guelte A, Wan L, Kang Y, Vazquez A, Gavard J, Arnoult D, Bidere N. The endoplasmic reticulum acts as a platform for ubiquitylated components of NF-kB signaling. Sci Signal. 2013 Sep 3 ;6(291).
-Soares F, Tattoli I, Arnoult D, Philpott DJ, Girardin SE. NLRX1 does not inhibit MAVS-dependent antiviral signaling. Innate Immun. 2013;19(4):438-48.
2012
-Zemirli N, Arnoult D. Mitochondrial anti-viral immunity. Int J Biochem Cell Biol. 2012 Sep;44(9):1473-6.
-Castanier C, Zemirli N, Portier A, Garcin D, Bidere N, Vazquez A, Arnoult D. MAVS ubiquitination by the E3 ligase TRIM25 and degradation by the proteasome is involved in type I Interferon production after activation of the antiviral RIG-I-Like receptors. BMC Biol. 2012 May 24;10(1):44.
2011
-Arnoult D, Soares F, Tattoli I, Girardin SE. Mitochondria in innate immunity. EMBO Rep. 2011 Sep 1;12(9):901-10.
-Edlich F, Banerjee S, Suzuki M, Cleland MM, Arnoult D, Wang C, Neutzner A, Tjandra N, Youle RJ. Bcl-x(L) Retrotranslocates Bax from the Mitochondria into the Cytosol. Cell. 2011 Apr 1;145(1):104-16.
-Castanier C, Arnoult D. Mitochondrial localization of viral proteins as a means to subvert host defense. Biochim Biophys Acta. 2011 Apr;1813(4):575-83.
2010
-Castanier C, Arnoult D. Mitochondrial dynamics during apoptosis. Med Sci (Paris). 2010 Oct;26(10):830-5.
-Chang CR, Manlandro CM, Arnoult D, Stadler J, Posey AE, Hill RB, Blackstone C. A lethal de novo mutation in the middle domain of the dynamin-related GTPase Drp1 impairs higher order assembly and mitochondrial division. J Biol Chem. 2010 Oct 15;285(42):32494-503.
-Castanier C, Garcin D, Vazquez A, Arnoult D. Mitochondrial dynamics regulate the RIG-I-like receptor antiviral pathway. EMBO Rep. 2010 Feb;11(2):133-8.
2009
-Arnoult D, Soares F, Tattoli I, Castanier C, Philpott DJ, Girardin SE. An N-terminal addressing sequence targets NLRX1 to the mitochondrial matrix. J Cell Sci. 2009 Sep 1;122(Pt 17):3161-8.
-Figueroa-Romero C, Iñiguez-Lluh JA, Stadler J, Chang CR, Arnoult D, Keller PJ, Hong Y, Blackstone C, Feldman EL. SUMOylation of the mitochondrial fission protein Drp1 occurs at multiple nonconsensus sites within the B domain and is linked to its activity cycle. FASEB J. 2009 Nov;23(11):3917-27
-Arnoult D, Carneiro L, Tattoli I, Girardin SE. The role of mitochondria in cellular defense against microbial infection. Semin Immunol. 2009 Aug;21(4):223-32.
-Bidère N, Ngo VN, Lee J, Collins C, Zheng L, Wan F, Davis RE, Lenz G, Anderson DE, Arnoult D, Vazquez A, Sakai K, Zhang J, Meng Z, Veenstra TD, Staudt LM, Lenardo MJ. Casein kinase 1alpha governs antigen-receptor-induced NF-kappaB activation and human lymphoma cell survival. Nature. 2009 Mar 5;458(7234):92-6.